
Neutrophils are weird (Part 1): rolling and squeezing
How neutrophils are made, how they move, and why they were missing from our datasets

This is part of a series on neutrophil biology. Neutrophils are the most abundant immune cell in the human body, but they have been historically underestimated. These posts cover what makes them unusual, from their production and mechanical behavior to their role in cancer.
If you look at any of the large single-cell atlases published in the last decade, you will notice that neutrophils, the most abundant immune cell in the human body, are often missing.
This is because single-cell RNA sequencing workflows are poorly suited for them. They require dissociating tissue into individual cells, and the mechanical forces involved destroy neutrophils before their RNA can be captured. This technical problem reinforced older biases that caused Neutrophils to be underestimated for a long time. They live for hours, not days, which makes them hard to study in culture. They were considered simple killers with a single job, so they attracted less research attention than T cells or macrophages. And when the big cell atlas projects arrived, the dissociation problem kept them out of the data entirely.
The result is that some of the most interesting aspects of neutrophil biology were discovered late, often using specialized techniques: microfluidic imaging, intravital microscopy, electron microscopy, and more recently, spatial transcriptomics. Image-based spatial transcriptomics methods like Xenium, CosMx or MERSCOPE are particularly promising for neutrophil research because they measure gene expression directly in intact tissue sections, without any dissociation step. This may cause a renaissance in neutrophil biology, one where we can finally study these cells at scale and in their native context. What these approaches reveal, taken together, is a cell that is genuinely unusual. Its production, lifespan, metabolism, mechanical behavior, and way of navigating blood vessels all work differently from what other immune cells do.
This post covers what neutrophils are, how they are produced, and how they get from the bloodstream to the site of infection. Each of these turns out to be more interesting than you would expect.
What makes neutrophils unusual
Neutrophils make up about 70% of all white blood cells in human blood. Their primary job is to find and destroy foreign entities: bacteria, fungi, and other pathogens. They can also attack cancer cells directly, although their relationship with cancer is complicated and will be the subject of a later post.
To maintain their numbers, the bone marrow produces roughly 1 billion neutrophils per kilogram of body weight per day. For a 70 kg person, that is about 70 billion neutrophils every 24 hours. During infections, this rate increases to 10 billion per kilogram per day.
Neutrophils live 12 to 24 hours. They arrive at the site of infection, deploy their weapons, and self-destruct, because a cell that kills indiscriminately needs to disappear quickly. The short lifespan prevents neutrophils from accumulating in tissue and causing the kind of sustained damage that leads to autoimmune disease. Compare this to memory T cells, which can persist for decades.
Their cytoplasm is packed with granules: intracellular compartments loaded with antimicrobial proteins and enzymes. We will return to how they use these later.
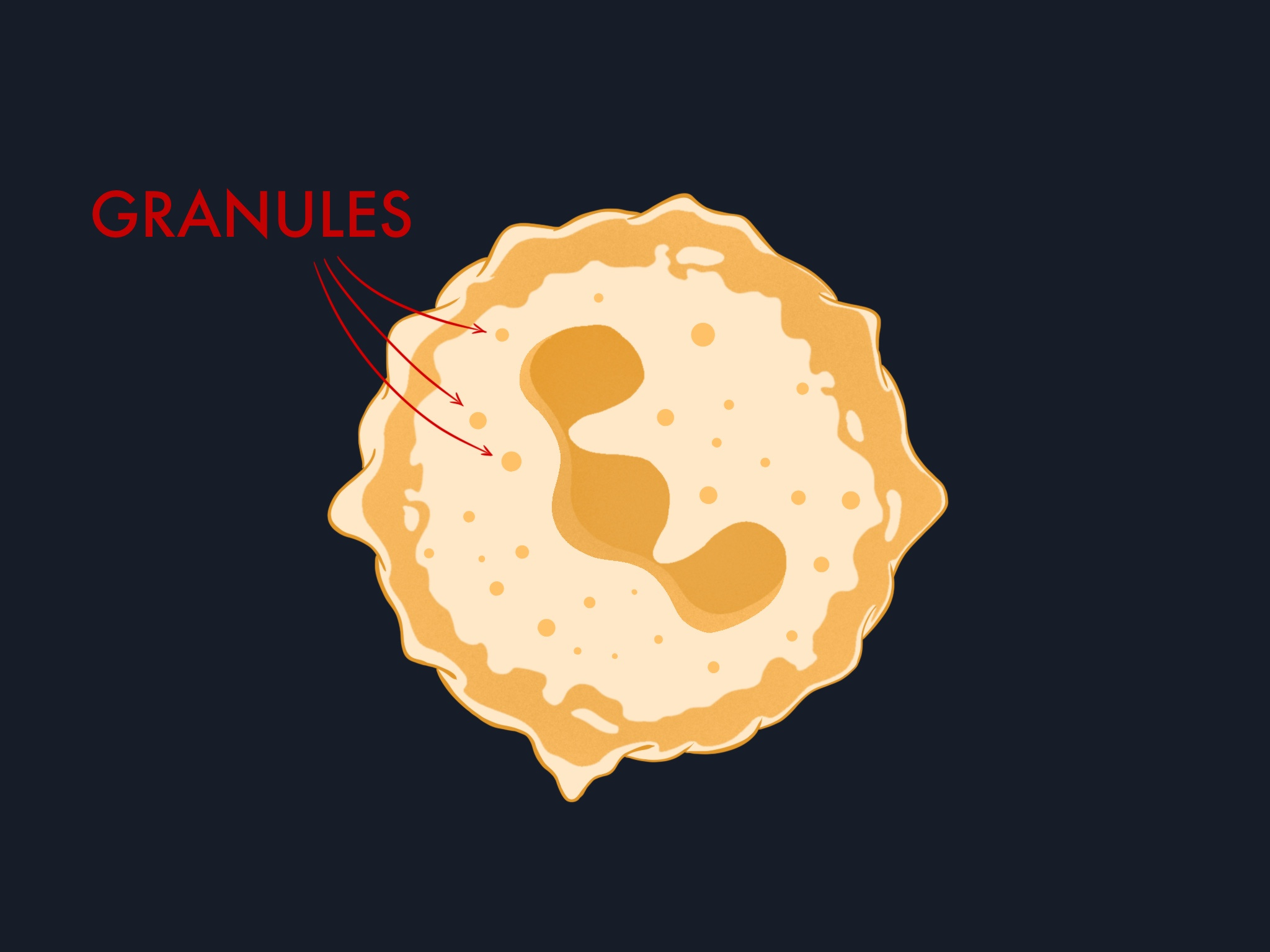
Their metabolism is also unusual. Most cells use oxygen primarily for energy production through mitochondrial respiration, but neutrophils have very few mitochondria. They rely on glycolysis for ATP and redirect oxygen toward a different purpose: fueling the enzyme NADPH oxidase, which generates toxic superoxide radicals to kill microbes.
How neutrophils reach the site of infection
Neutrophils circulate in the bloodstream until they receive signals directing them toward damaged or infected tissue. The mechanism that guides them is chemotaxis: they detect concentration gradients of signaling molecules called chemokines and move toward increasing concentrations.
In the 1950s, David Rogers at Vanderbilt University filmed one of the clearest demonstrations of this. His time-lapse shows a single neutrophil on a blood smear, weaving between red blood cells to chase down a Staphylococcus aureusbacterium. The neutrophil tracks the bacterium through several changes of direction, catches it, and engulfs it.
But getting to a site of infection in a living body involves more than following a gradient. Neutrophils circulate in the bloodstream at high speed and need to exit through the walls of blood vessels at the right location. This exit follows a four-step adhesion cascade.
The four-step cascade: rolling, arrest, crawling, transmigration
Rolling. When the endothelial cells lining a blood vessel detect nearby inflammation, they display selectin molecules on their surface. Circulating neutrophils bind these selectins loosely, which slows them from full blood flow speed to a controlled roll along the vessel wall. This rolling gives neutrophils time to scan the endothelial surface for stronger activation signals.
Arrest. As rolling neutrophils encounter higher concentrations of chemokines displayed on the endothelial surface, integrins on the neutrophil switch to a high-affinity state. The cell stops rolling and firmly attaches to the vessel wall.
Crawling. The arrested neutrophil crawls along the inner surface of the blood vessel, following the chemokine gradient to locate the best exit point, where the signal is strongest.
Transmigration. The neutrophil squeezes between endothelial cells and crosses the vessel wall into the tissue. This step requires the neutrophil to deform its entire body through gaps only a few micrometers wide.
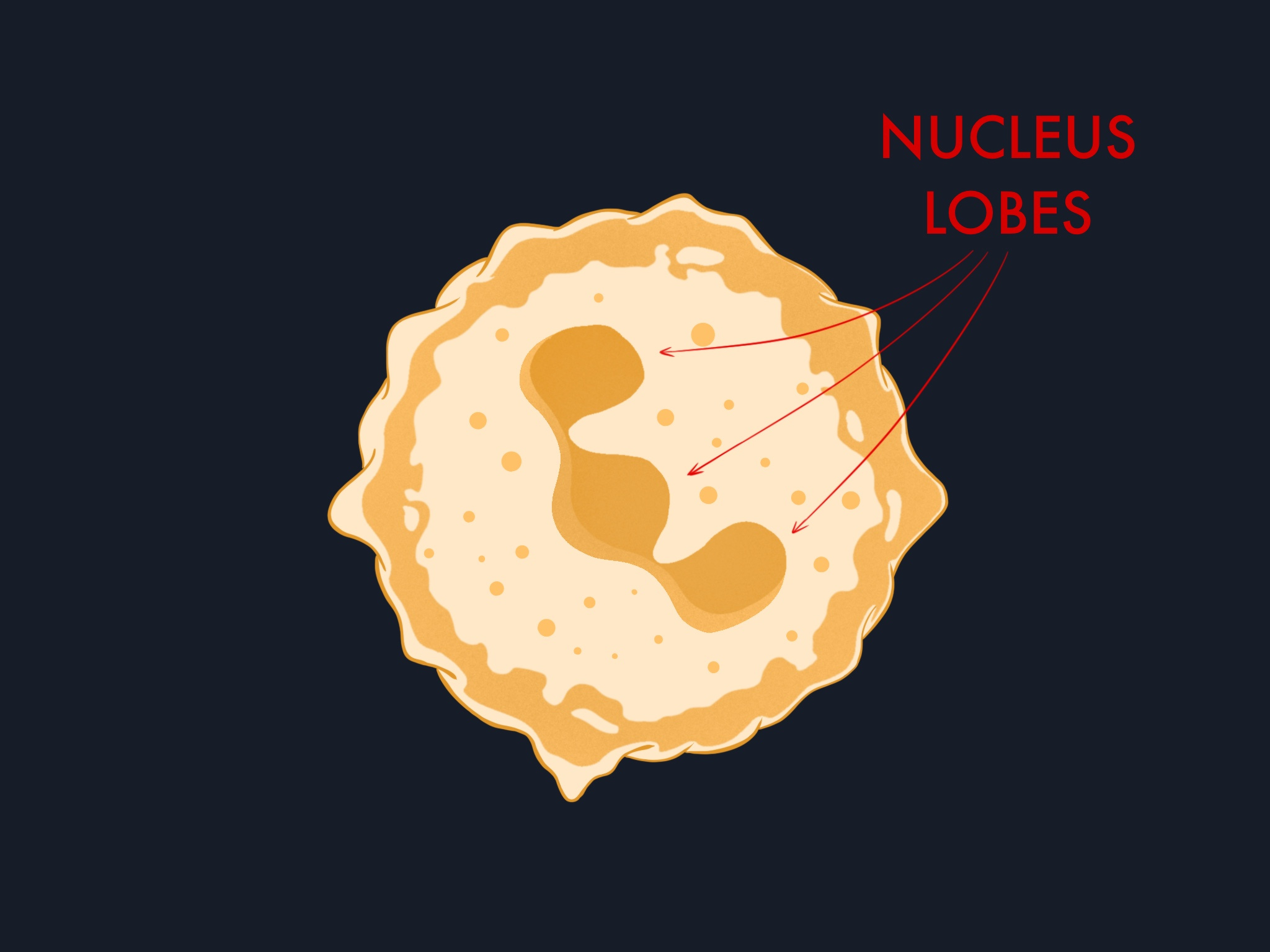
Other leukocytes also exit blood vessels, but most depend on the endothelium actively widening its junctions to let them through during transmigration. Neutrophils have a structural adaptation that reduces this dependency. Their nucleus is multi-lobulated, shaped like a series of connected lobes rather than a single sphere. This makes it highly deformable, allowing neutrophils to squeeze through tighter gaps.
Tethers and slings: how neutrophils roll under extreme flow
Rolling, the first step of the cascade, presents a physical problem. During inflammation, blood flow through small vessels increases, and the resulting shear forces would strip most leukocytes off the vessel wall. Neutrophils can maintain stable rolling at shear stress levels 10 times higher than what other leukocytes tolerate. How they manage this was unclear until 2012.
Using a technique called quantitative dynamic footprinting (qDF), Prithu Sundd and colleagues at the La Jolla Institute observed something unexpected. As a rolling neutrophil is pulled forward by blood flow, the drag force stretches the cell membrane at the back into a thin tube called a tether. These tethers can extend up to 40 micrometers, and they act as anchors that slow the cell down.
The new finding was what happens next. When a tether detaches, it does not retract into the cell as previously assumed. Instead, it persists and swings forward, wrapping around the front of the rolling neutrophil. Sundd’s group named these structures slings. Not all tethers become slings, but those that do create a self-made adhesive surface for the cell to roll on.
Slings are covered in adhesion molecules arranged in patches. As the neutrophil rolls forward and the sling wraps around it, these patches engage with the vessel wall one by one. When one patch fails under force, the next picks up the load almost immediately. This stepwise peeling keeps the cell anchored far more effectively than a single tether attachment. Two slings and two tethers together can balance more than 75% of the forward force acting on a rolling neutrophil.
The slings also carry molecules that bind to proteins on the neutrophil’s own surface, pulling the sling tighter around the cell body and further reducing rolling speed. The cell wraps itself in its own braking system.
Conclusion
Neutrophils are built for speed and access. They roll under shear forces that would strip other cells off the vessel wall. They deploy tethers and slings to stay attached. They squeeze through endothelial gaps with a nucleus shaped for deformation. All of this happens before they reach the site of infection, and before they have used any of their killing mechanisms.
In the next post, we will look at what happens after neutrophils arrive in tissue: how the physical act of crossing a barrier activates their killing capacity, and the three antimicrobial mechanisms they deploy once they reach their target.
Neat!